

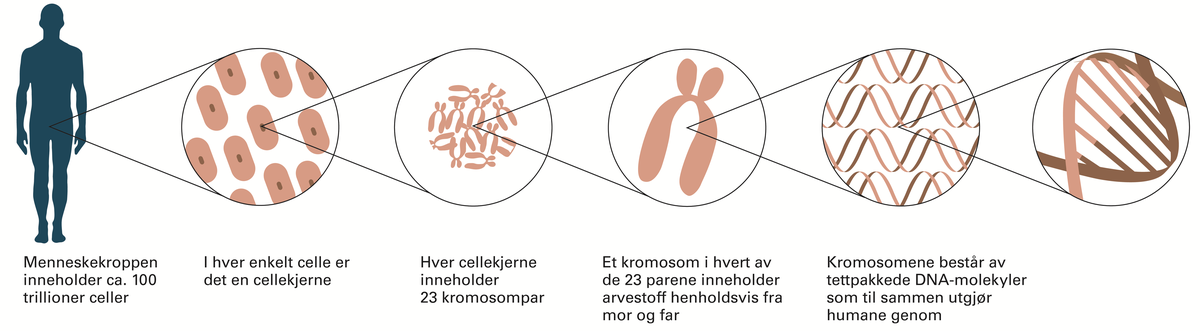
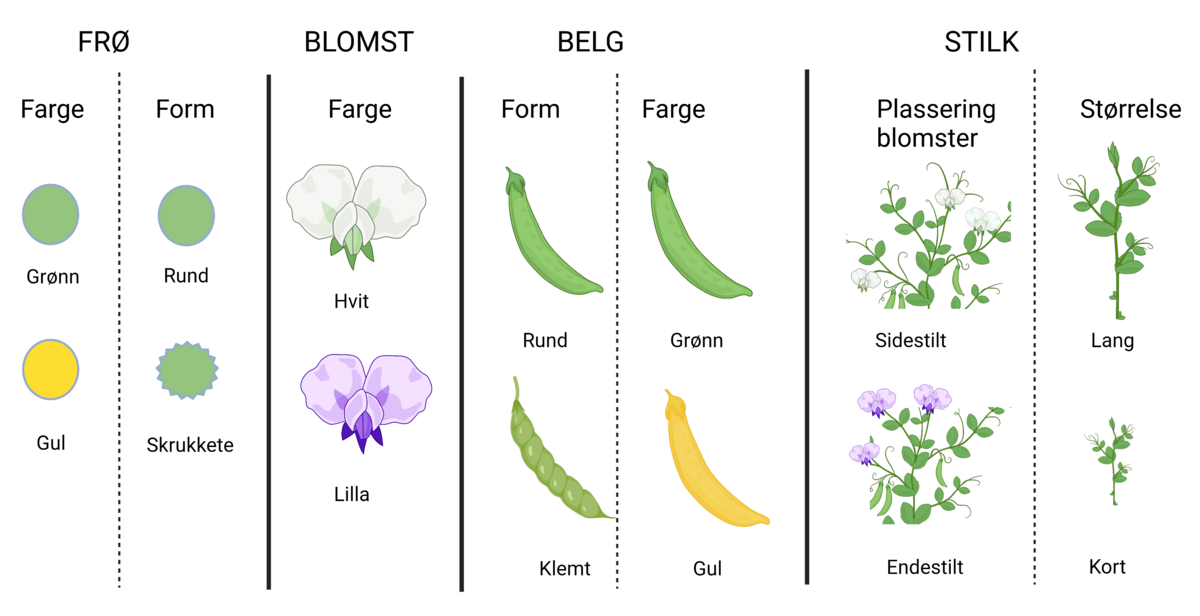
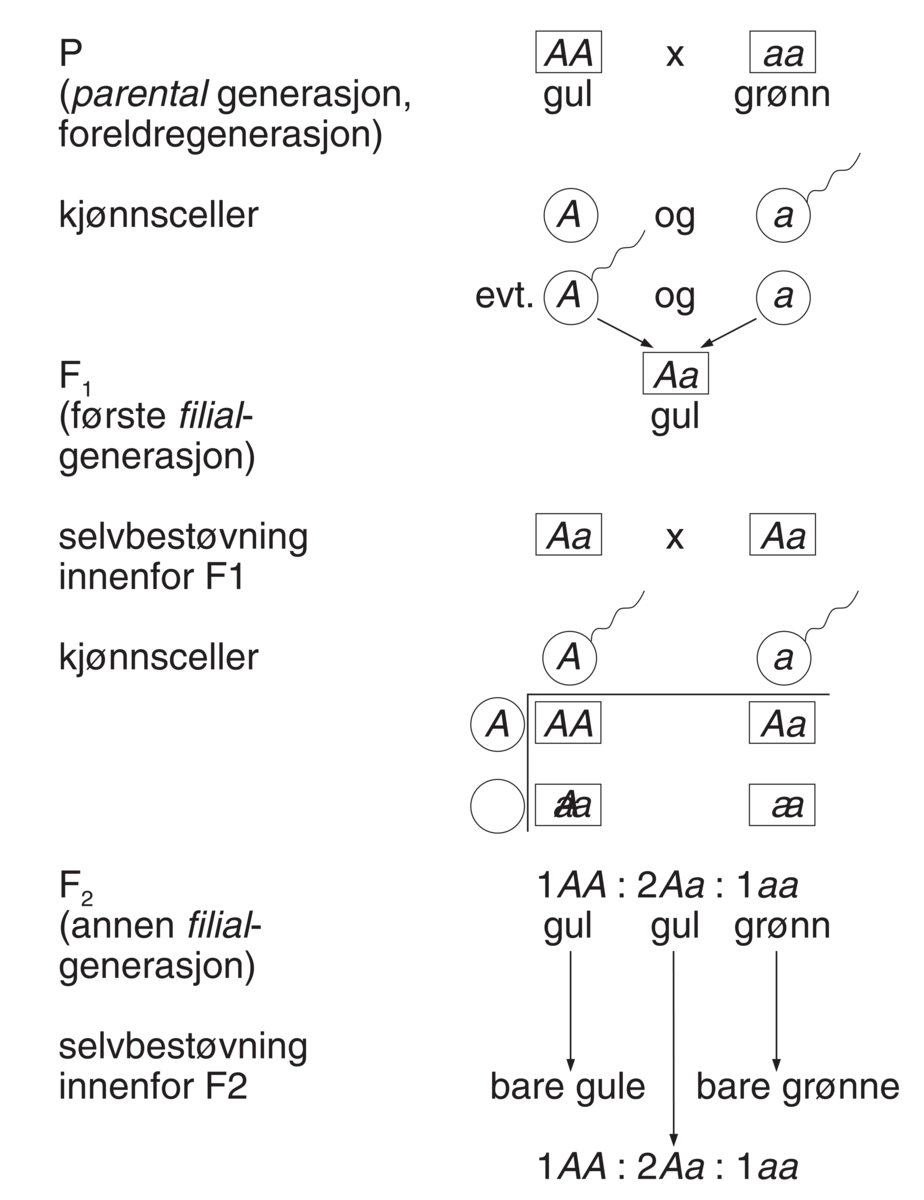
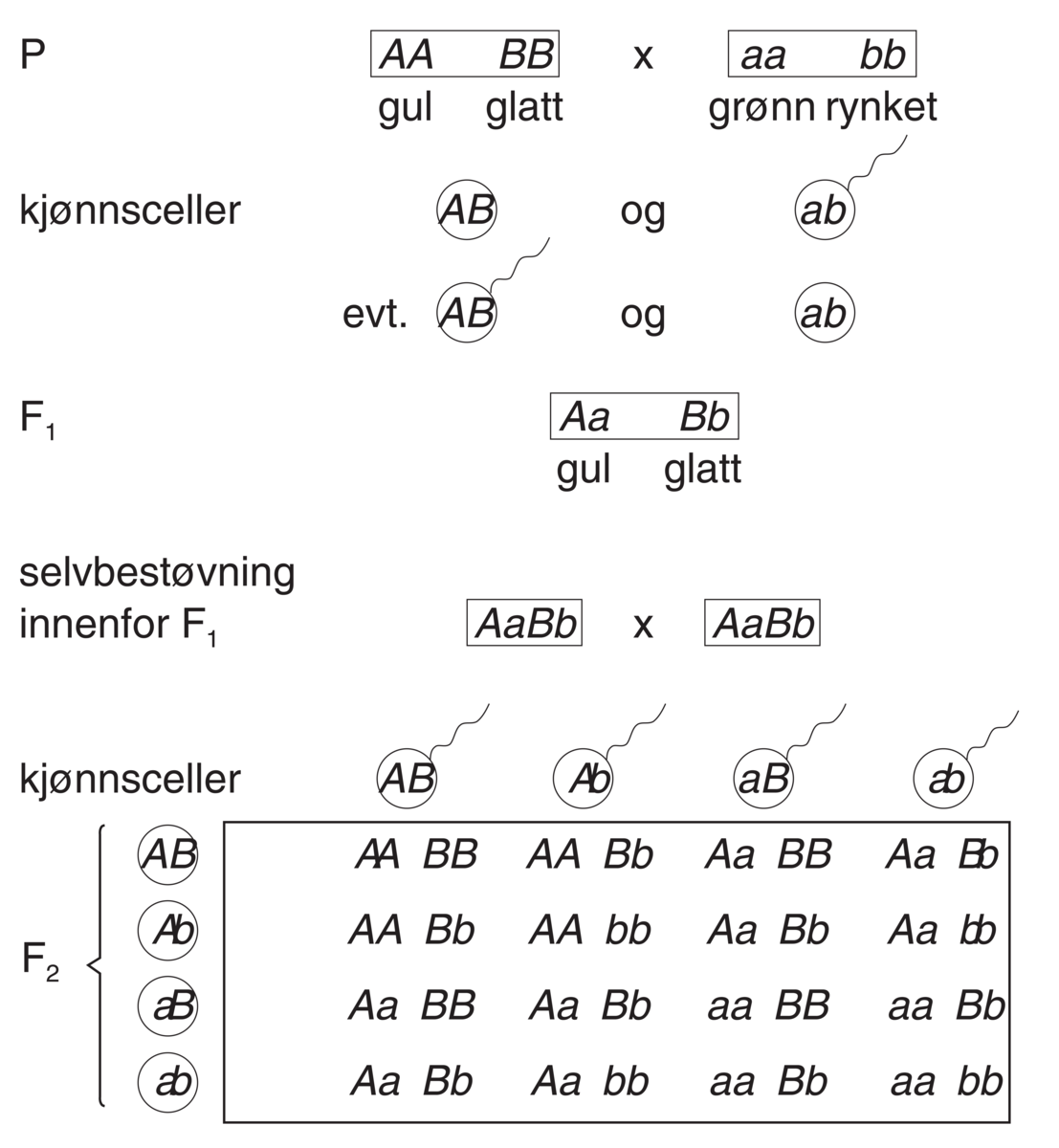
Genetikk er et fag i biologien hvor man studerer gener, genenes variasjon og hvordan genene arves fra én generasjon til den neste. Gener, også kalt arveanlegg, er oppskrifter for egenskaper hos levende organismer slik som fargen på en blomst, at insekter har seks ben, høyden på et tre, hårfarge, og mange andre ting. Genetikken studerer alle typer egenskaper, både de som varierer innen samme art og de som er forskjellige fra art til art.
Faktaboks
- Uttale
- genetˈikk
- Etymologi
-
avledet av gresk γενετικός genetikos 'genitiv/generativ' som igjen kommer fra γένεσις 'genesis' 'opprinnelse, tilblivelse'
- Også kjent som
-
arv, arvelære
Genetikken ligger til grunn for mange grener av moderne biologi, som studier av slektskap (systematikk) og evolusjon, og i tillegg har det stor relevans innen medisin (medisinsk genetikk).
Menneskets genetikk ble kartlagt i human genomprosjektet (HGP).


Kommentarer
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.