
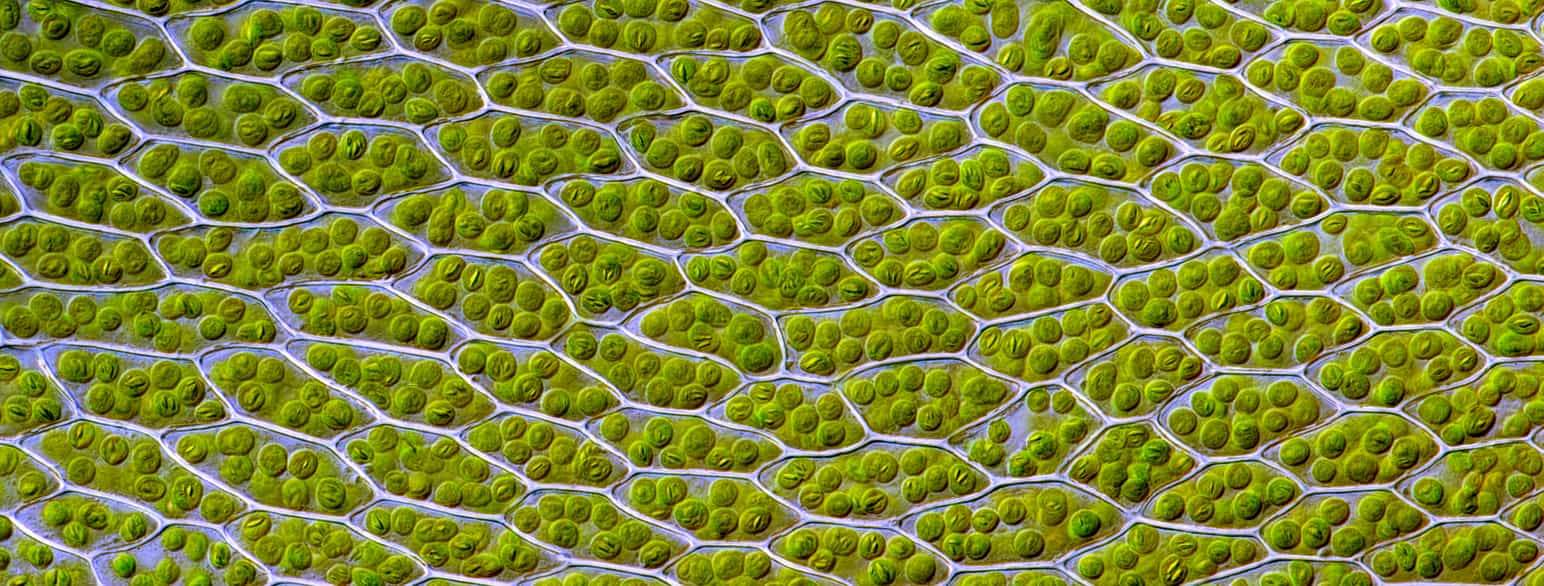
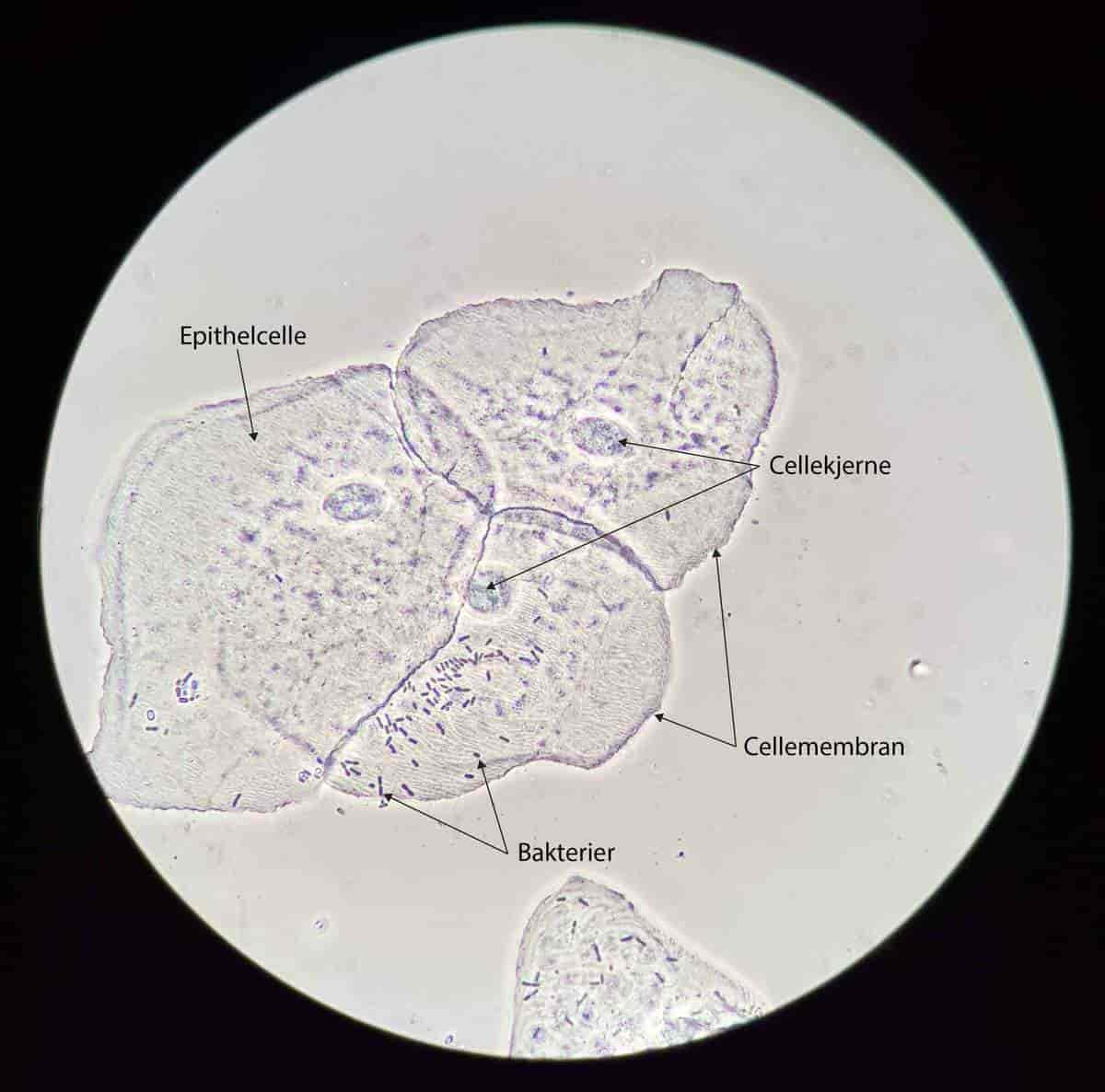
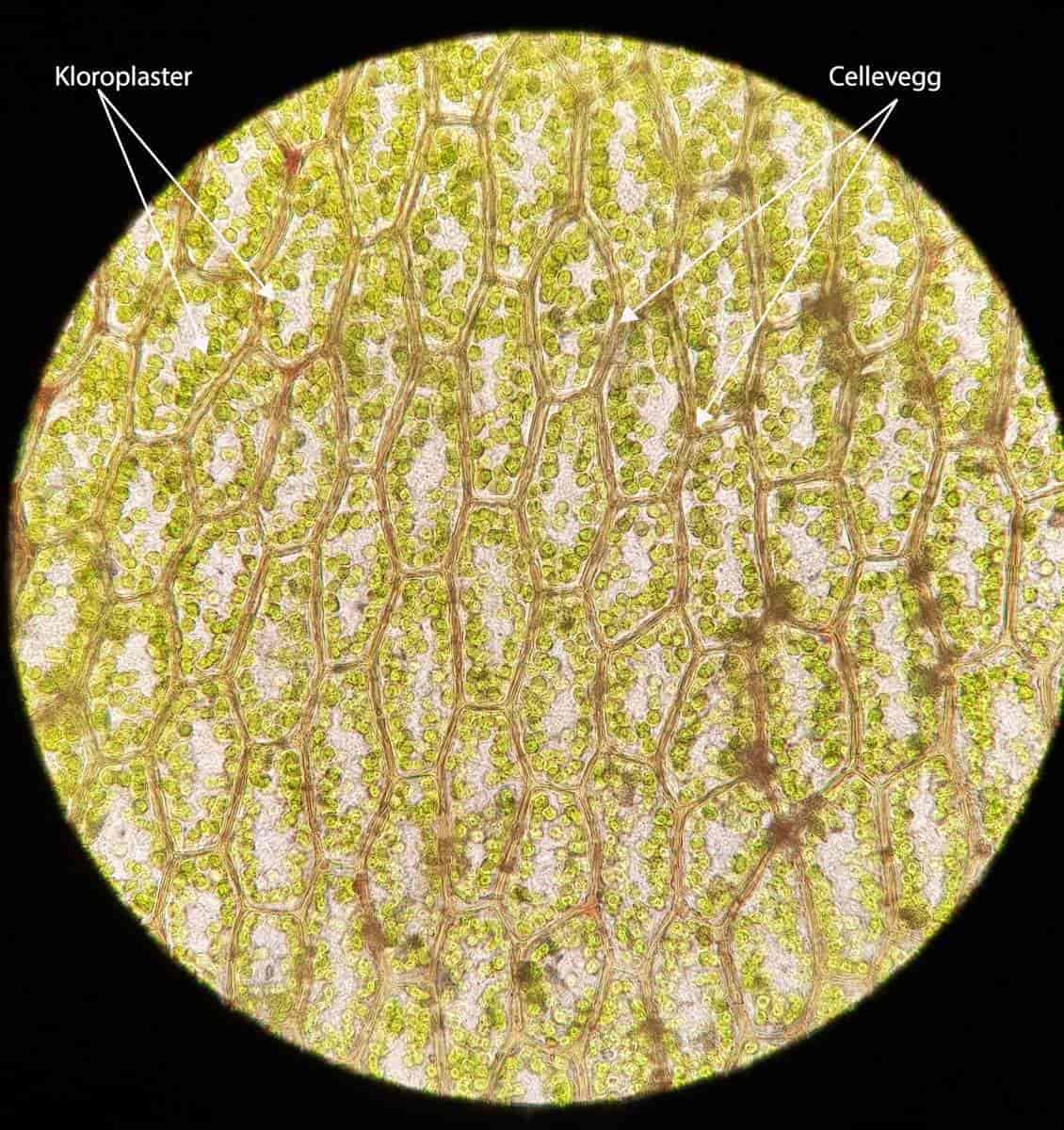
En celle er den minste enheten for liv. Alle organismer er bygget opp av enten én eller flere celler.
Alle bakterier og noen dyr, planter og sopp består av bare én celle. Disse kalles encellede organismer. Noen organismer kan bestå av flere identiske celler og danner da kolonier. De aller fleste organismer består av mange ulike celler, som samarbeider på ulike måter. Antallet celler i slike organismer er vanligvis enormt stort, og avhenger blant annet av hvor stor organismen er. En elefant har for eksempel flere celler enn en mus. Hvert menneske består av et sted mellom 30 og 40 billioner celler.
De fleste celler er så små at man ikke kan se dem med øynene, men noen er veldig store, som et egg. Noen celler (nerveceller) kan strekke seg helt fra én ende av kroppen til en annen. De fleste celler kan vi kun observere gjennom mikroskop.
Alle celler har en hinne av fett som fungerer som en grense til omgivelsene og til andre celler, en cellemembran.
Alle celler har noen egenskaper til felles, men det er også mye som kan gjøre celler veldig ulike. Ulike typer celler kan løse ulike oppgaver, som å være celler i muskler, celler som beskytter mot sykdom, eller celler som sprer sykdom. Ulike celler i en organisme har ulike funksjoner og i en organisme har vi derfor mange ulike celletyper. For eksempel har mennesker omtrent 220 ulike celletyper i kroppen. Nesten alle celler inneholder arvemateriale som forteller hva cellen skal gjøre.
Alle celler kan dele seg ved hjelp av celledeling og på denne måten videreføres arvematerialet til neste generasjon celler. Alle celler stammer fra eksisterende celler. Dette innebærer at alle celler stammer fra en urcelle som var begynnelsen på alt liv på Jorda for omtrent tre milliarder år siden.
Alle celler trenger også energi for å utføre ulike oppgaver. Celler er derfor i stand til å omdanne næringsstoffer til energi i en prosess som kalles celleånding.
Alle celler har en bestemt levetid. Noen celler lever i noen få timer andre i flere år. Celledød er en nøye kontrollert prosess både blant encellede og flercellede organismer. Når cellen dør fyller den ikke lenger kriteriene for liv. Døde celler løses etter hvert opp til ulike organiske og uorganiske molekyler som gjenbrukes av andre organismer.


{kind=link}
Kommentarer (2)
skrev Tor Stian Holte
I Sender, Ron, Fuchs, Shai & Milo, Ron 2016. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLOS Biology https://doi.org/10.1371/journal.pbio.1002533 oppgis tallet på celler i menneskekropphen til 30 billioner. Har dere andre eller nyere kilder?
svarte Halvard Hiis
Hei! Fagansvarlig har nettopp oppdatert artikkelen. Hun henviser til at nyere publikasjoner ofte viser til 37 billioner, jamfør følgende artikkel: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6085824/ Vennlig hilsen Halvard i redaksjonen
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.