
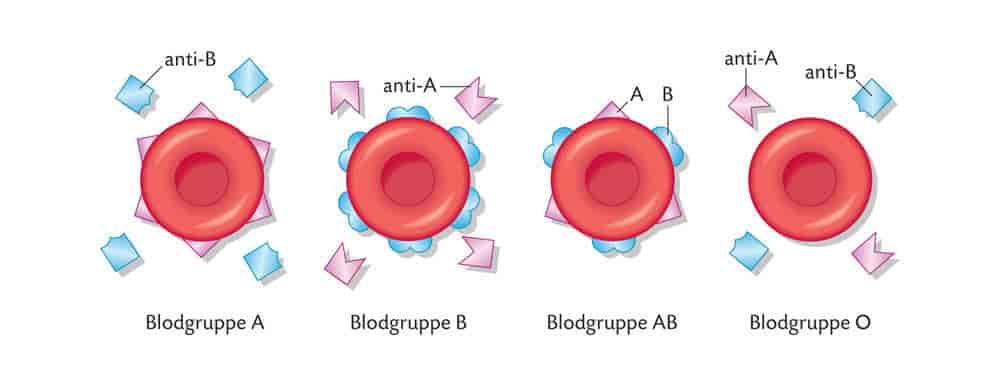
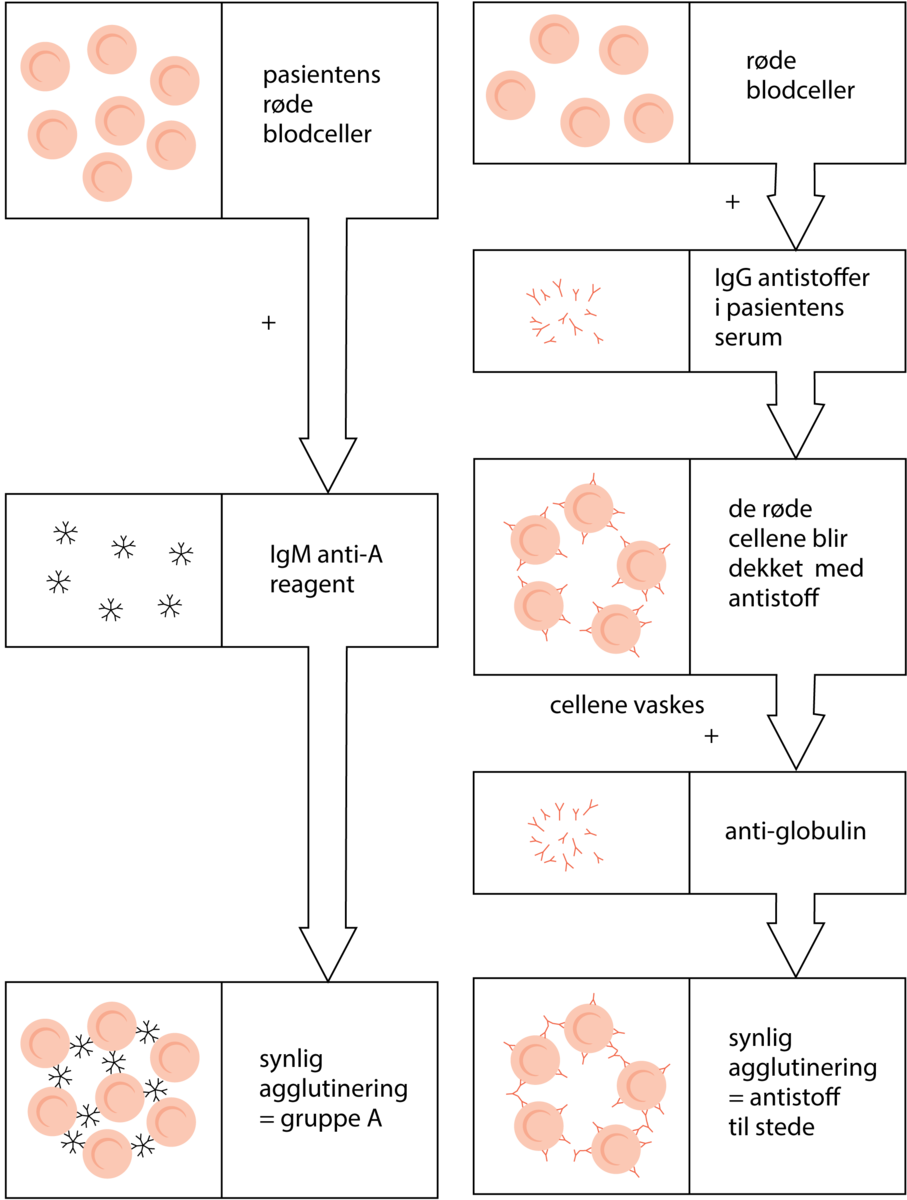
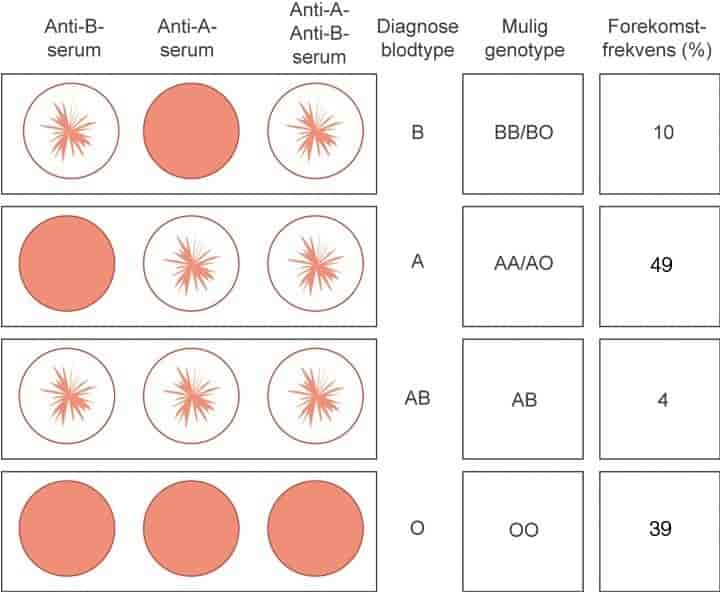
Blodtyper er måter å kategorisere blod på grunnlag av små forskjeller i blodcellene. De røde blodcellene hos mennesker har små variasjoner fra individ til individ i en del proteiner og karbohydrater i cellemembranen. Mange av disse forskjellene kan utløse immunreaksjoner hos mottakeren hvis de kommer inn i et annet menneske. Forskjeller som kan utløse immunreaksjoner kalles for blodtyper. Blodtyper defineres altså ved sin reaksjon med spesifikke antistoffer.
Blodtyper kan deles inn i ulike blodtypesystemer. De viktigste blodtypesystemene er ABO-systemet og Rh-systemet.
Blodtypeantigener og -antistoffer har helsemessig betydning bare ved blodoverføring, transplantasjon og svangerskap.

Kommentarer
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.