
Respirasjonssenteret er en samlebetegnelse på flere grupper av nevroner som til sammen styrer lungeventilasjonens rytme og intensitet.
respirasjonssenteret

Respirasjonssenteret. Aktivitetsprofilen til respiratoriske nevroner i løpet av 1,5 (en og en halv) pustesyklus. De respiratoriske nevroner er identifisert på grunnlag av deres aktivitet under inspirasjons- og ekspirasjonsfasen og det responsmønster de dermed tildeler nervus phrenicus, som innerverer den viktigste inspirasjonsmuskelen, diafragma.
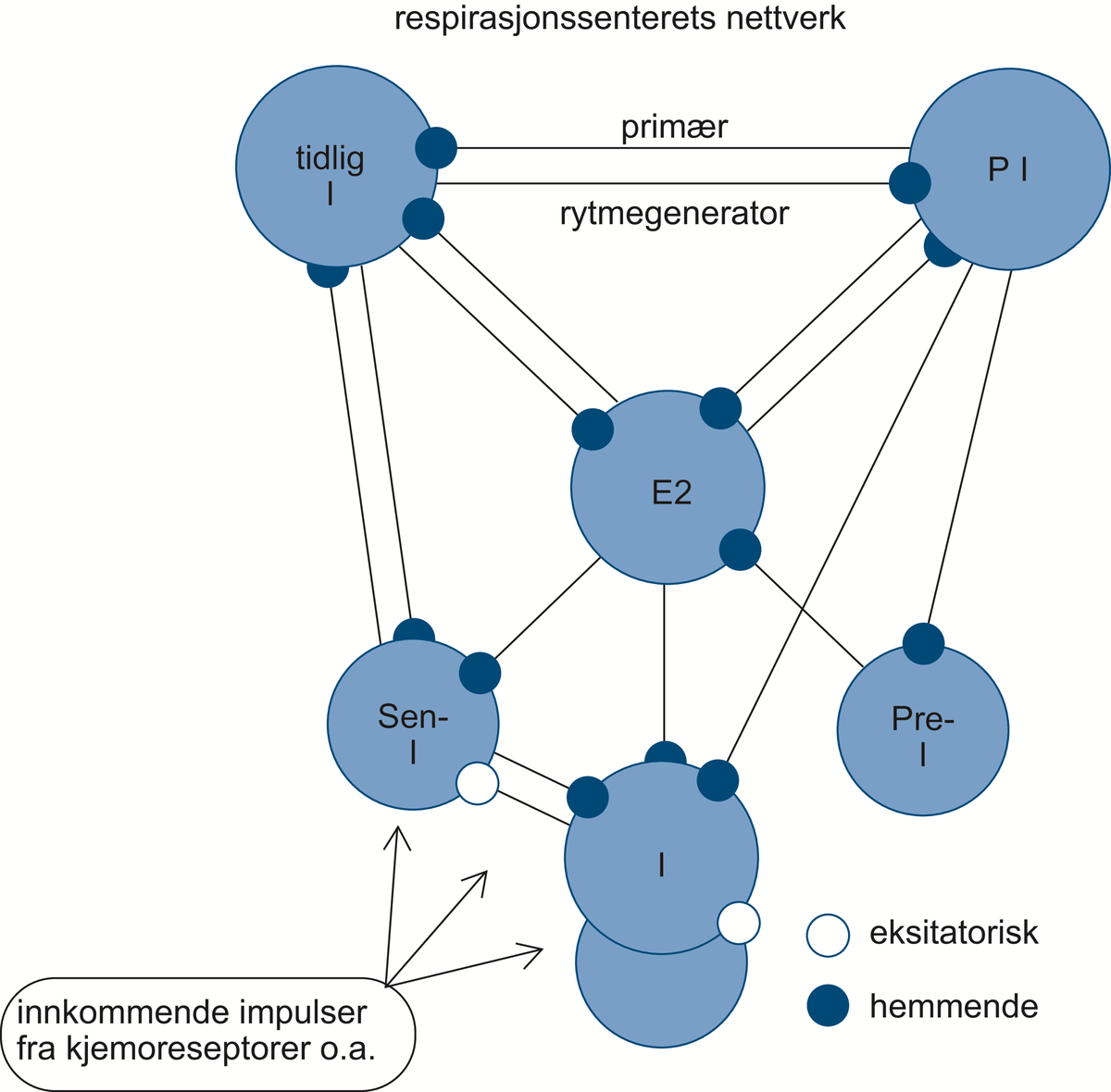
Respirasjonssenteret. Diagrammet under viser hvordan nevrongruppene gjensidig påvirker hverandre gjennom synaptiske forbindelser, dels hemmende, dels eksiterende. Den primære rytmegenerator utgjøres av «tidlig-inspiratoriske» (Tidlig I) og «post-inspiratoriske» (PI)-nevroner.
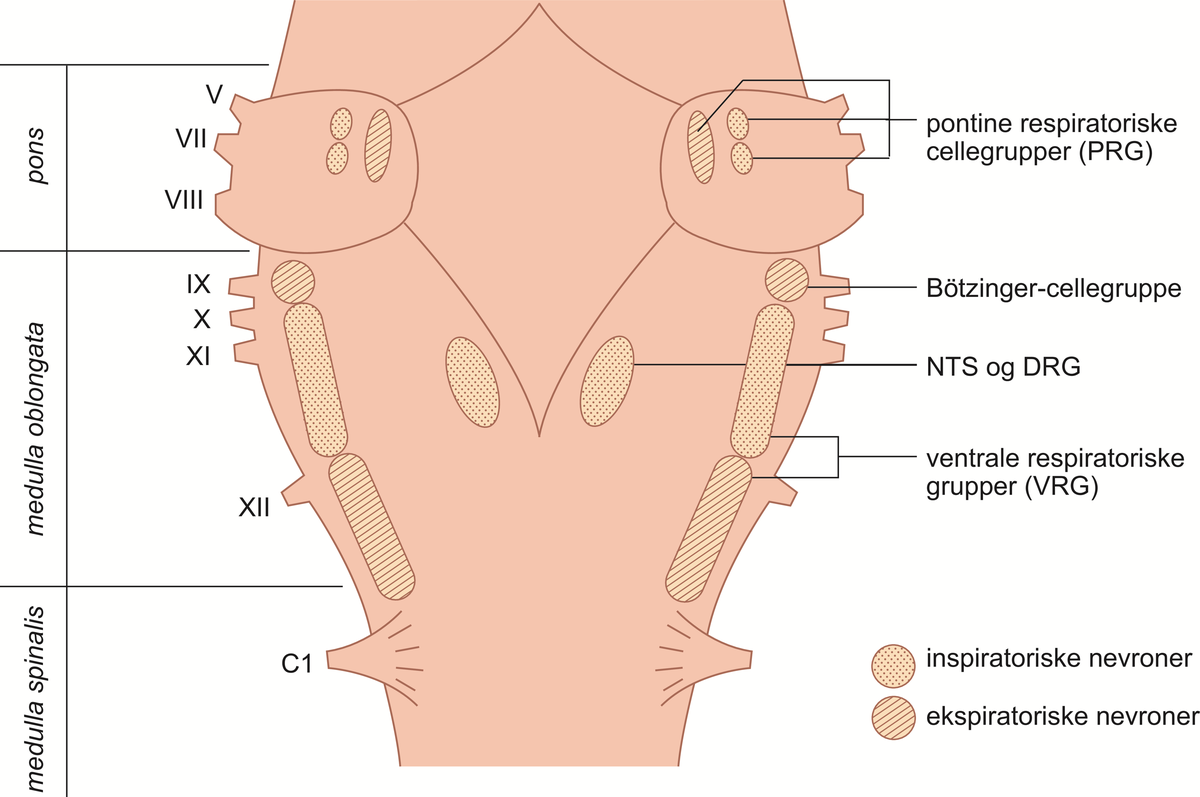
Respirasjonssenteret. Lokalisering i hjernen av de nevrongrupper som til sammen utgjør respirasjonssenteret. PRG: pontine respiratoriske gruppe; DRG: dorsale respiratoriske gruppe; NTS: nucleus tractus solitarius. Romertallene angir utløpssteder for de tilsvarende hjernenerver.
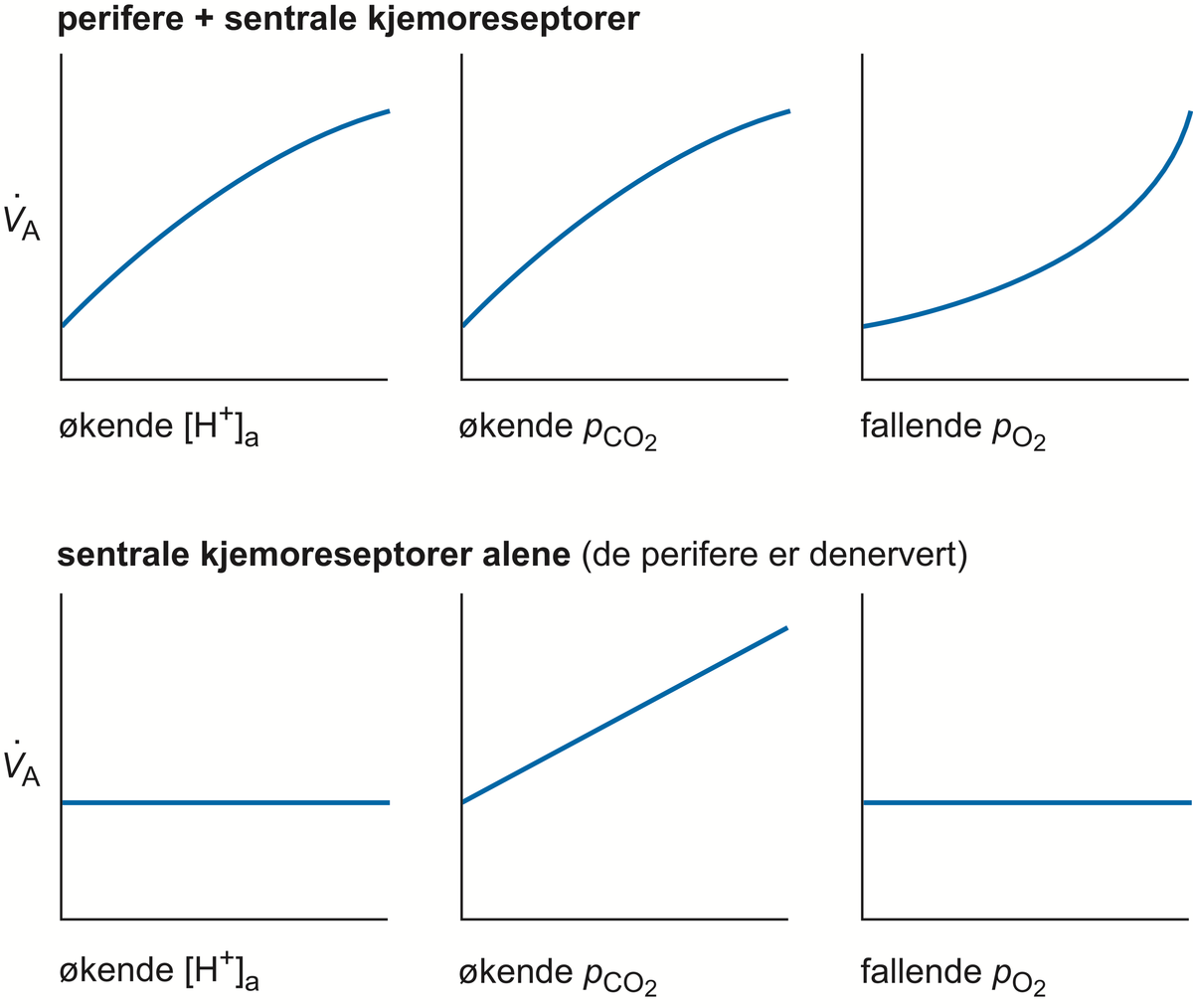
Respirasjonssenteret. Skjematisk ventilasjonsrespons ved enkeltvis økning av hydrogenionekonsentrasjonen [H+] og karbondioksidtrykket pCO2, og ved reduksjon av pO2 i arterielt blod (a). Den øverste rekken viser responsene hos et intakt individ. I den nedre rekken er de perifere kjemoreseptorene denervert (ikke fungerende). Nå er det bare pCO2-økning som gir ventilasjonsstimulering. A er alveolær ventilasjon vol/tid. VA = alveoleventilasjon.
Lokalisering
I den forlengede marg (medulla oblongata) finner man bilateralt (tosidig) tre slike nevrongrupper: den dorsale respiratoriske gruppe (DRG), den ventrale respiratoriske gruppe (VRG) og Bötzingers kompleks. I nedre del av pons (hjernebroen) finner man, også bilateralt, den pontine respiratoriske gruppe (PRG).
Funksjon
Disse nevrongruppene står i kontakt med hverandre gjennom nervebaner som til dels er hemmende, dels eksiterende, og som gjennom en slik nettverkskobling sørger for optimal dybde og frekvens av pustebevegelsene. Tilbakekoblingssignaler fra strekkreseptorer i lungene, fra mekanoreseptorer i muskler, sener og ledd, og fra sentralnervøse og arterielle kjemoreseptorer, gir nødvendig informasjon til respirasjonssenteret slik at en korrekt alveoleventilasjon kan opprettholdes under vekslende forhold, fra dyp søvn til maksimalt muskelarbeid. Derved kan pO2, pCO2 og pH holdes nær konstant i arterielt blod og i cerebrospinalvæsken.
Respirasjonssenteret integrerer de mange innkommende signalene og sender rytmisk utadførende (efferente) nerveimpulser til de motoriske forhornscellene som styrer kontraksjonen av de forskjellige respirasjonsmusklene. Lungeventilasjonen påvirkes også av stimuli som smerte og temperaturforandringer og viljebestemte handlinger. Irritasjonsstimuli i neseslimhinne og bronkialvegg, og også akutt lungeødem, kan via egne autonome refleksbaner forbigående endre pustemønsteret fullstendig og gi hurtig overfladisk ventilasjon, eventuelt nysing og hoste.
Det er ikke avklart om det i respirasjonssenteret er celler med pacemaker-egenskaper, eller om den basale pusterytmen utelukkende er et resultat av egenskaper ved det nevronale nettverket.
Inndeling
Detaljerte undersøkelser de senere år har vist at nervecellene i respirasjonssenteret kan inndeles i seks funksjonelle typer vurdert etter deres nerveimpulsaktivitet («fyringsprofil») mot nervus phrenicus som innerverer den største inspirasjonsmuskelen, diafragma (mellomgulvet). Tre av de seks hovedtypene nevroner er aktive under inspirasjon. Det er de tidliginspiratoriske (tidlig-I) nevronene, som er aktive i første del av inspirasjonen, de inspiratoriske nevronene (hele-I), som er aktive under hele inspirasjonen, og de sent inspiratoriske (sen-I) nevronene, som er aktive i siste del av inspirasjonen. De postinspiratoriske nevronene (PI), derimot, er aktive under første del av ekspirasjonen. De vil kunne bremse utstrømning av luft og derved øke ekspirasjonsfasens varighet. Ekspiratoriske nevroner (E2) er aktive under det meste av ekspirasjonen, mens nervus phrenicus (som går til en inspirasjonsmuskel) er passiv. Preinspiratoriske (pre-I) nevroner fyrer helt mot slutten av ekspirasjonen og litt over i starten av neste inspirasjon.
Respirasjonssenterets cellegrupper står i resiprok (gjensidig) kontakt med andre cellegrupper i hjernestammen, som regulerer hjertets minuttvolum og den perifere karmotstanden. Ut fra funksjonelle betraktninger er dette en rimelig ordning: Lungeventilasjonen utgjør første etappe i oksygentransporten fra atmosfæren og til de enkelte cellene i kroppen. Kretsløpet utgjør neste etappe.
De sentrale kjemoreseptorene befinner seg også i hjernestammen, anterolateralt ved utløpet av niende og tiende hjernenerve, men er anatomisk adskilt fra respirasjonssenteret. Disse reseptorene stimuleres av en økning i pCO2 og/eller et fall i pH i mellomcellevæsken og cerebrospinalvæsken. En surere cerebrospinalvæske vil aktivere disse cellene som så, i sin tur, øker impulstrafikken til respirasjonssenterets nevroner. Resultatet vil bli økt lungeventilasjon, større utlufting av CO 2 og normalisering av pH.
Under muskelarbeid vil nerveimpulser fra reseptorer i de arbeidende musklers sener, og fra ledd, nå frem til respirasjonssenteret via nervebaner i den forlengede margs bakhorn, og stimulere til økt lungeventilasjon. Det er også mulig at det foreligger et såkalt motorisk program på høyere nivå i sentralnervesystemet, som aktiveres under muskelarbeid. Man mangler fortsatt en del kunnskaper om hvordan disse viktige systemene er regulert.

Kommentarer
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.