
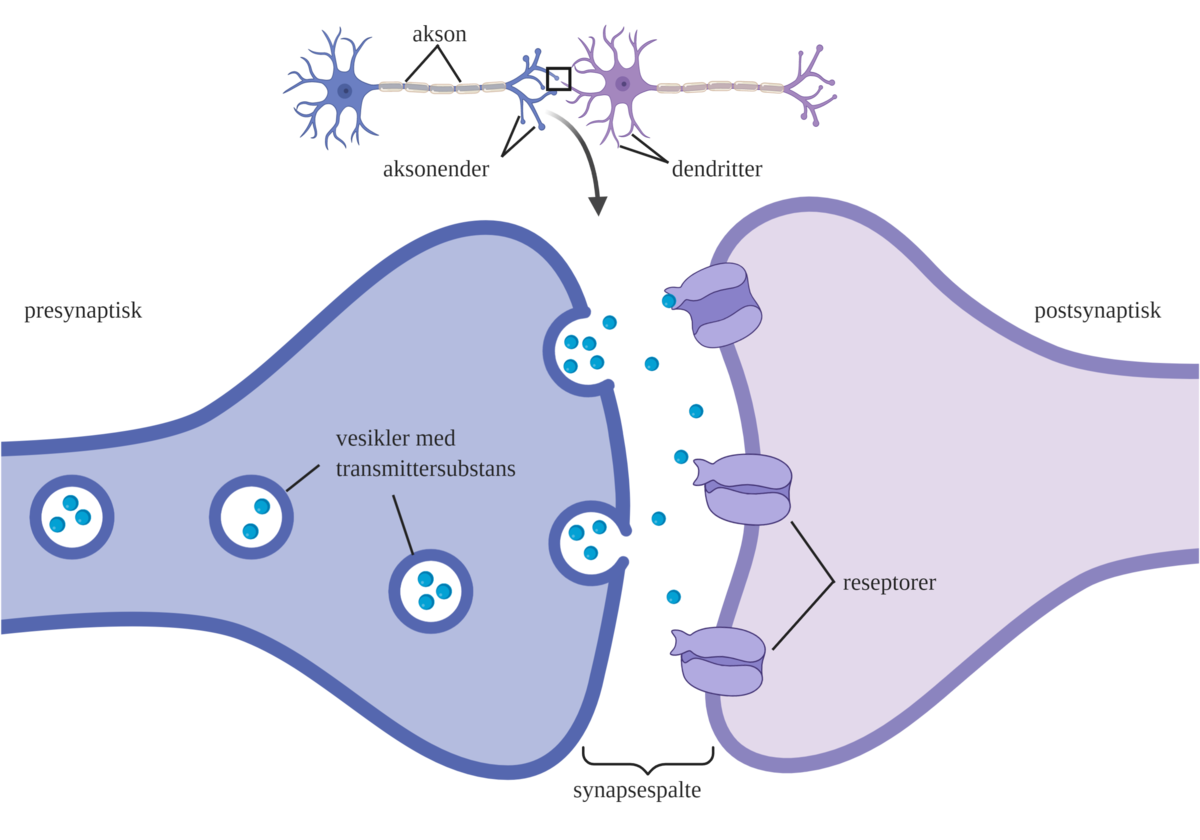
Synapse er et kontaktsted mellom en nervecelle og en annen nervecelle, en muskelcelle eller en kjertelcelle.
Cellen som sender signalet har lagret signalstoffer i små blærer. Når den blir stimulert vil den slippe ut signalstoffene i den lille spalten mellom de to cellene (synapsespalten). Noen vanlige signalstoffer er acetylkolin og noradrenalin.
Signalstoffene flyter over til mottakercellen og binder seg til reseptorer der. Reseptorene kan være ionekanaler eller metabotrope reseptorer. Når et signalstoff binder seg til en reseptor kan cellen bli aktivert (eksitert) eller hemmet (inhibert). I synapsespalten er det enzymer som bryter ned signalstoffet og dermed «skrur av» signalet.
Mange av legemidlene som brukes i behandlingen av nevrologiske og psykiske sykdommer påvirker signalformidlingen over synapser på en eller annen måte.
Kommentarer
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.