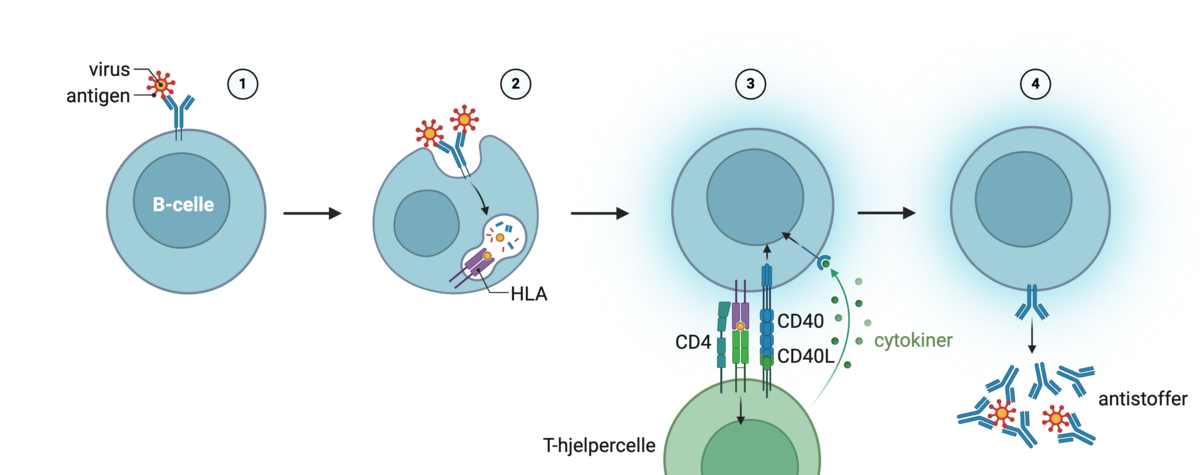
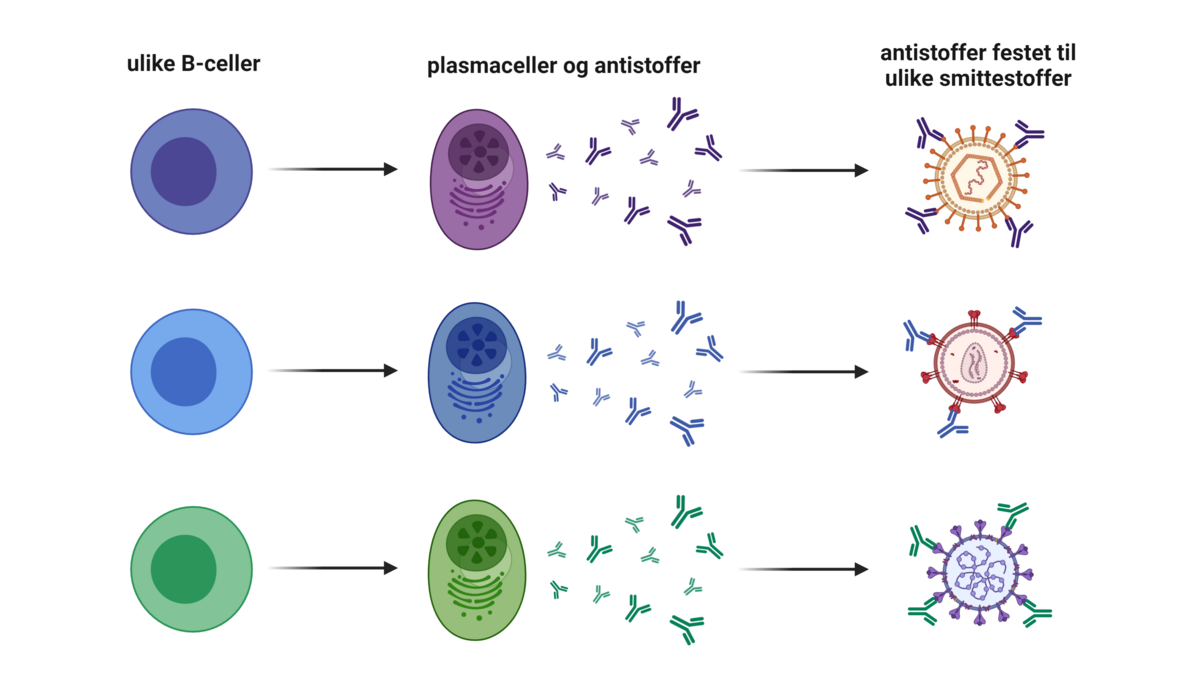
Plasmaceller produserer antistoffer. Antistoffer har samme struktur som den originale B-cellereseptoren, men er ikke lenger festet til cellemembranen. Fellesbegrepet for B-cellereseptorer og antistoffer er «immunglobuliner». B-cellereseptorer kan kalles «membranbundne immunglobuliner». Antistoffer kan kalles «løselige immunglobuliner», ettersom de ikke sitter fast i noe. Antistoff slippes ut av plasmacellene og kan ikke gå inn i celler på egen hånd. Det betyr at de kan binde til ting utenfor celler, det vil si i plasma og i vevsvæske.
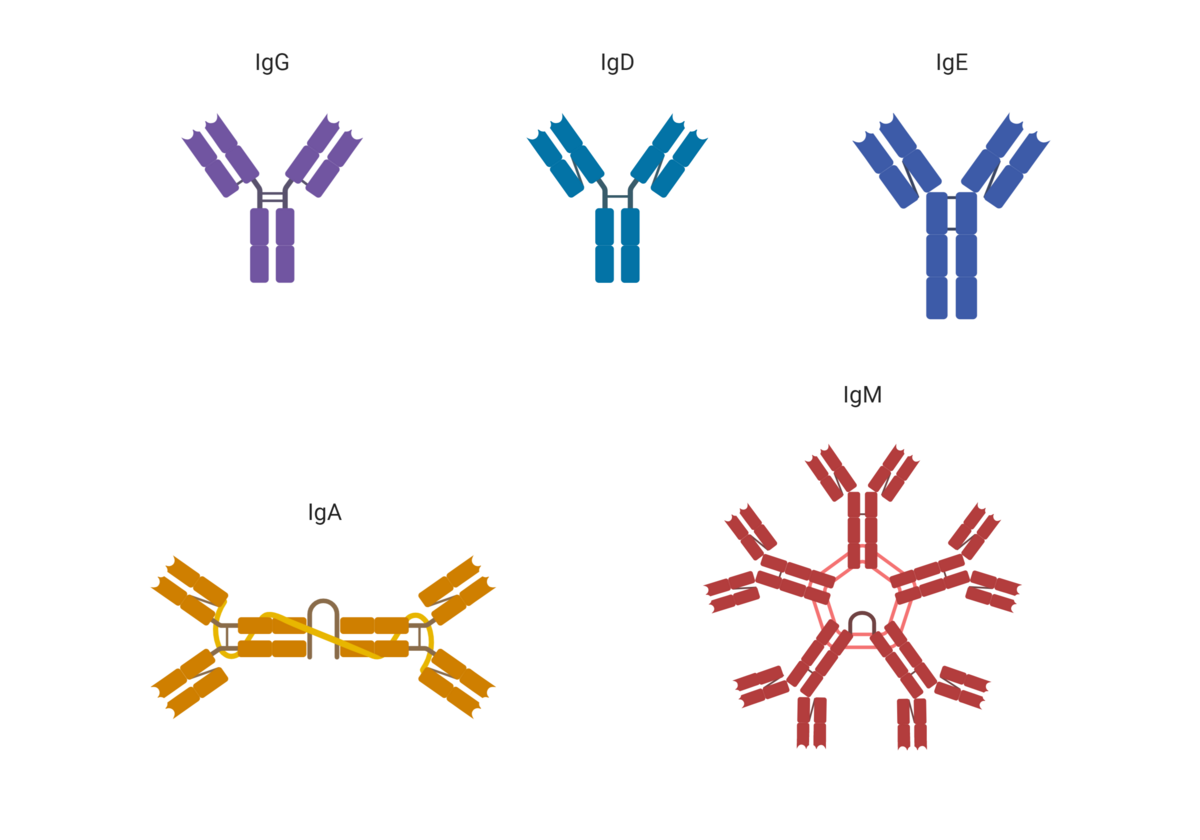
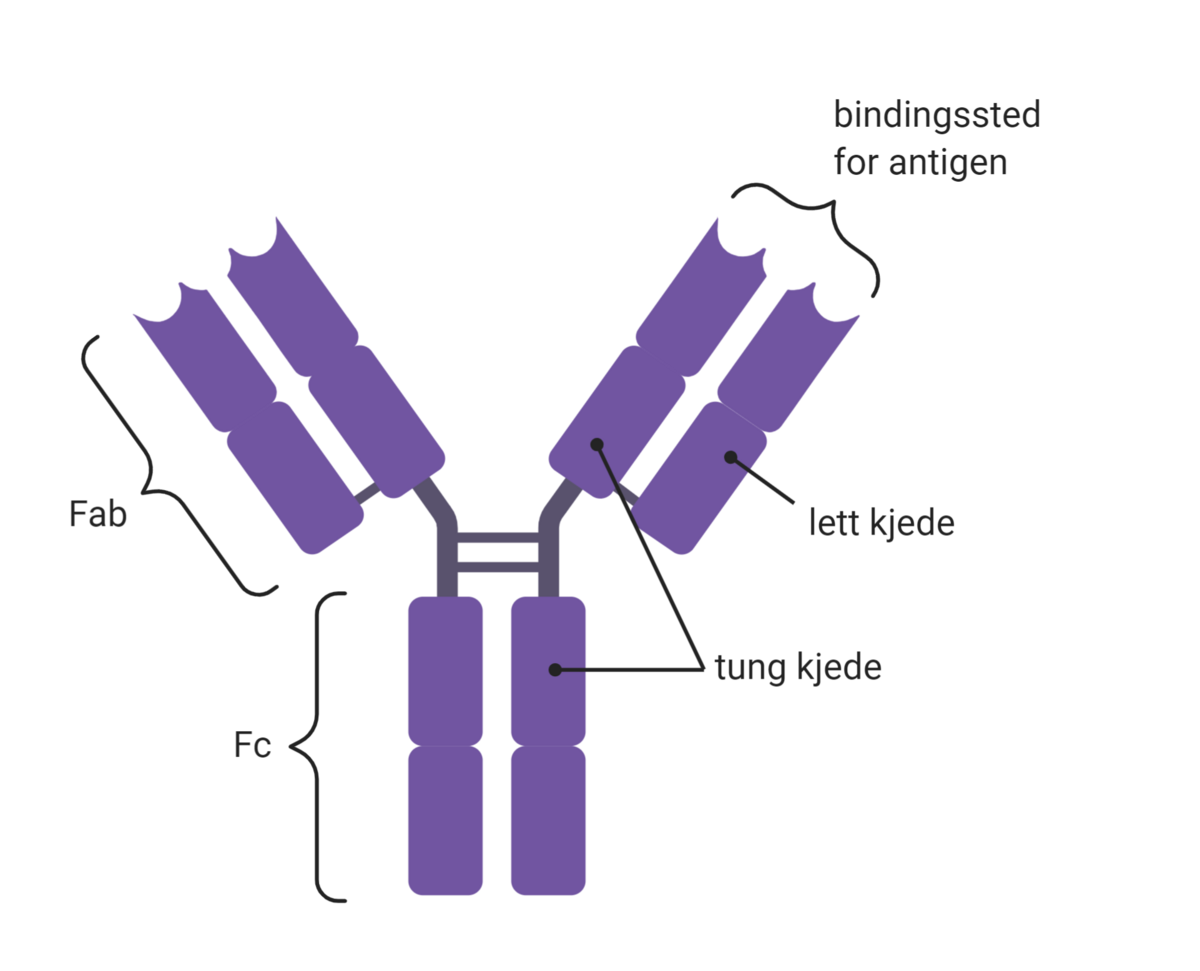
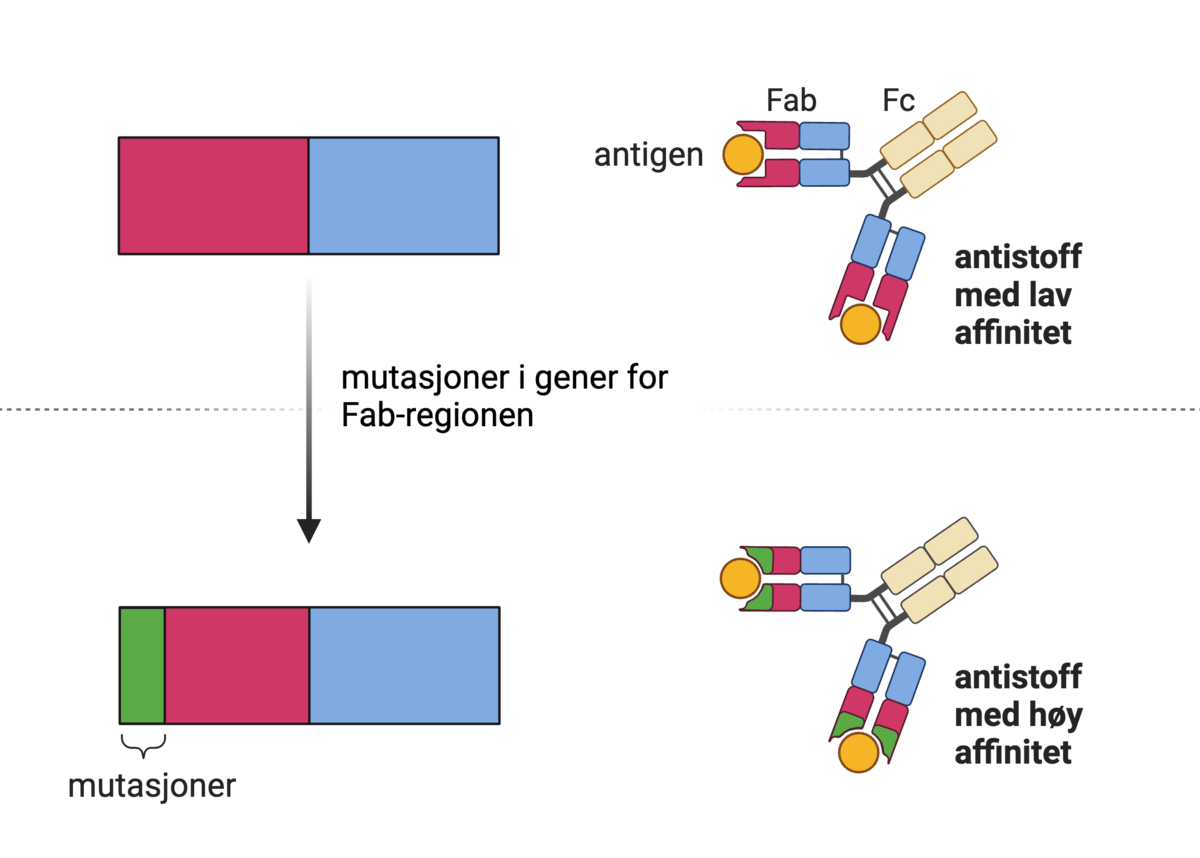
Strukturen til et antistoff kalles immunglobulin-monomer. Den består av to Fab-regioner og én Fc-region. Fab-regionene binder antigen. Fc-regionen kan binde til effektorsystemer. Med «effektorsystemer» menes andre deler av immunsystemet, som makrofager, granulocytter eller komplementsystemet. Antistoffer vil derfor fungere som et bindeledd mellom antigen og immunsystemet. Ved å bringe disse sammen, økes sannsynligheten for at effektorsystemene dreper og fjerner smittestoffet. Antistoffer kan også binde til giftstoffer (toksiner) og virus utenfor celler, og forhindre at disse kan binde og skade kroppens celler.
Det finnes fem hovedklasser av antistoffer: IgA, IgD, IgE, IgG og IgM. Disse har ulik Fc-region. Noen immunceller har reseptorer som kan binde Fc-regionen til IgG. Andre typer immunceller har reseptorer som kan binde Fc-regionen til IgE. Ulike antistoffklasser kan dermed aktivere forskjellige effektorsystemer, og derfor gi opphav til ulike typer immunresponsen. Hvilken antistoffklasse som er den beste i en gitt situasjon, avhenger av hva slags type infeksjon man er utsatt for.
Antistoffene kan sirkulere i blodet, eller de kan slippes ut på en slimhinne.


Kommentarer
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.