
Lymfocytter er en gruppe hvite blodceller som spiller en avgjørende rolle i immunsystemet. De er blant annet viktige i kampens forsvar mot smittestoff, som bakterier og virus.
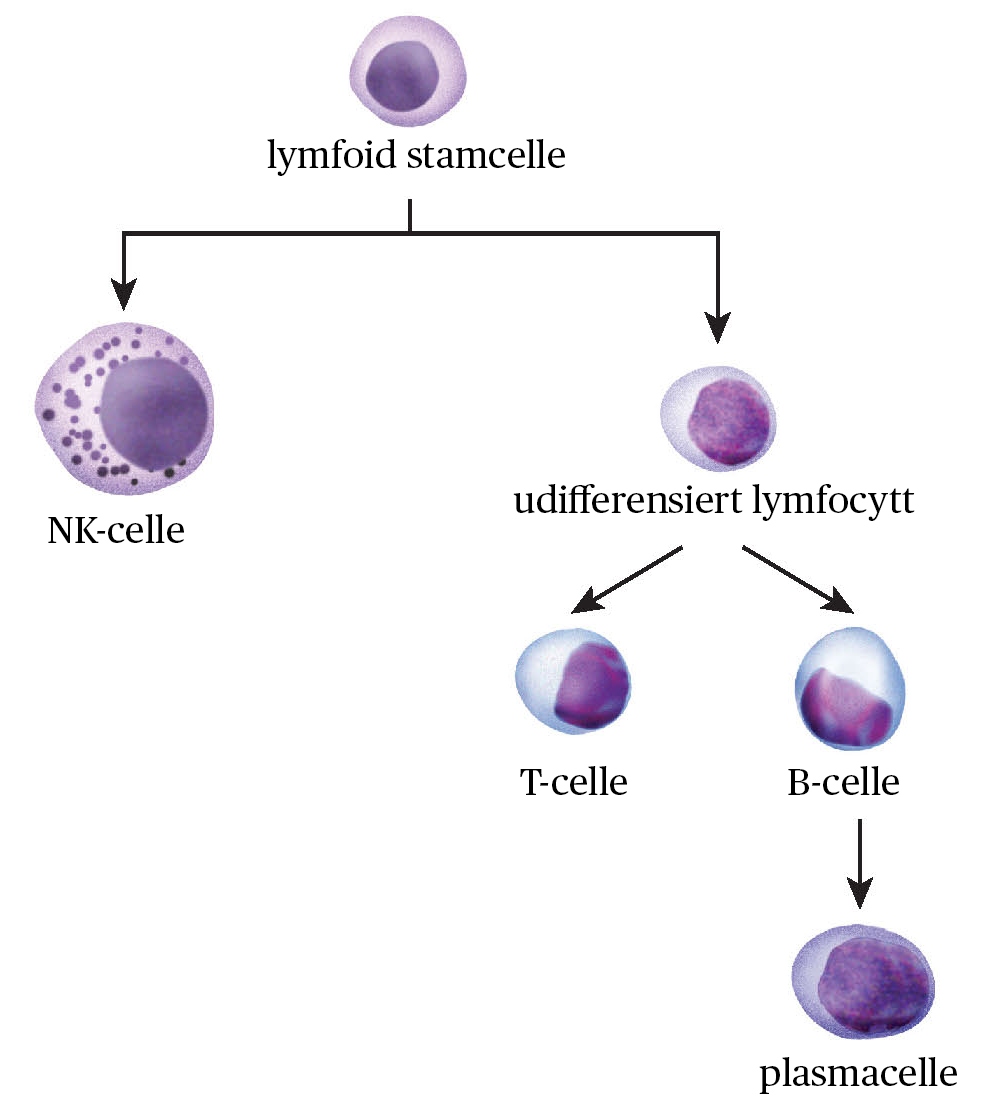
Det er to hovedtyper lymfocytter: B-celler og T-celler. Disse cellene tilhører det ervervede immunsystemet. Det finnes også en gruppe lymfocytter i det medfødte immunsystemet. Den viktigste typen her kalles naturlige dreperceller (NK-celler, natural killer cells).
Lymfocyttene er små og kulerunde, og de har en liten, mørk cellekjerne. Lymfocyttene utgjør om lag én tredjedel av de hvite blodcellene i blodet.
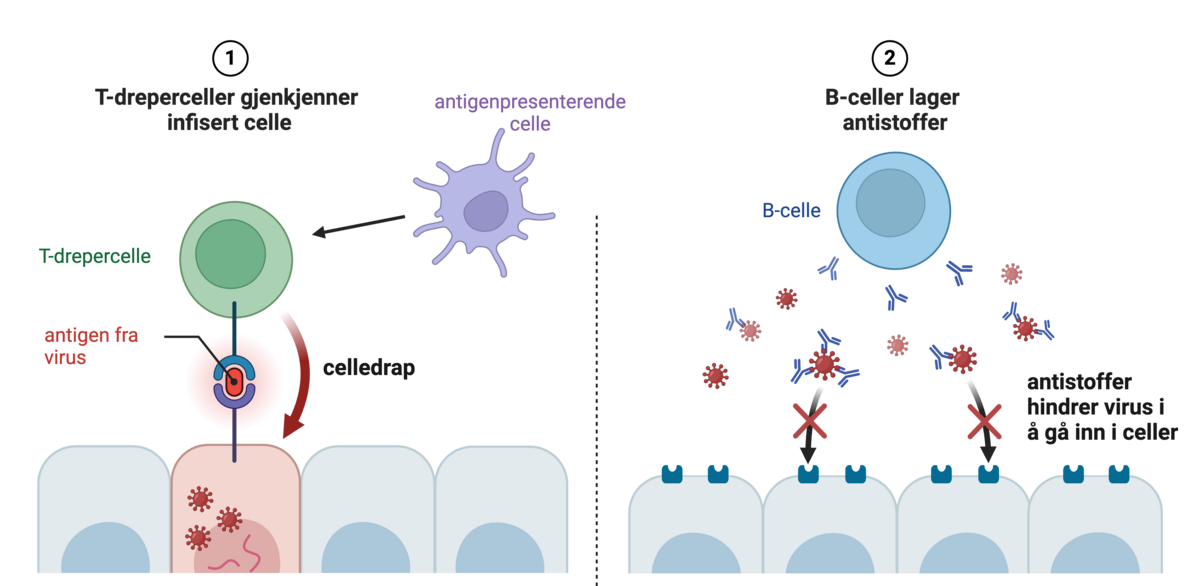
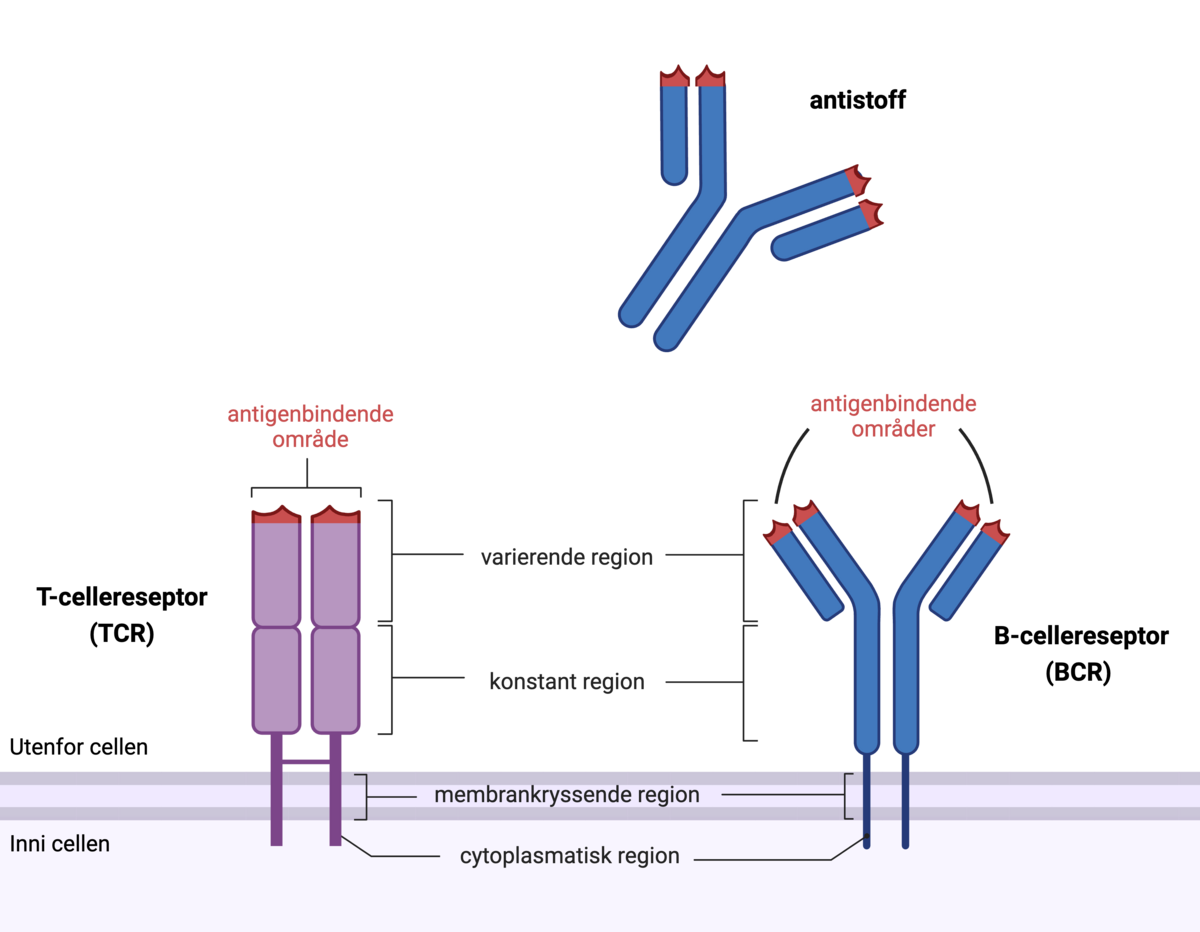
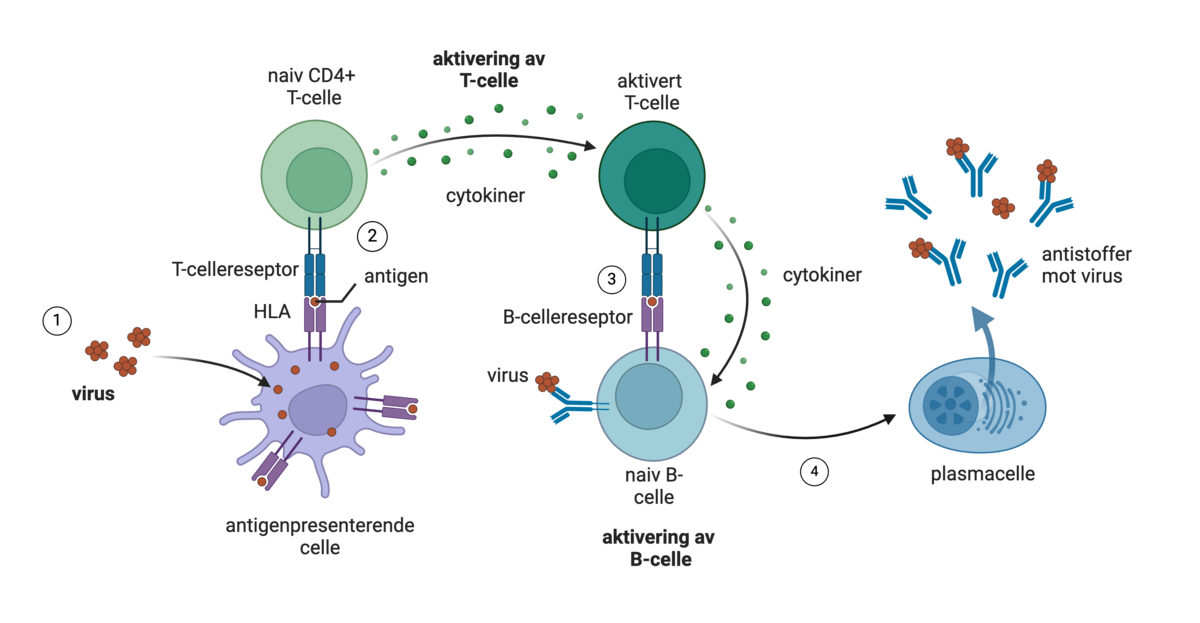
T-celler og B-celler er celler i immunforsvaret som har helt ulike roller. T-celler kan gjenkjenne celler i kroppen som er angrepet av virus. B-celler kan lage antistoffer som kan stanse virusene før de kommer inn i celler.


Kommentarer
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.